PLK1 (基因名), Serine/threonine-protein kinase PLK1 (蛋白名), plk1_bovin.
产品名称:
Bovine PLK1/ Serine/threonine-protein kinase PLK1 ELISA Kit
丝氨酸/苏氨酸蛋白激酶PLK1
货号:
E1662b
商标:
EIAab®
监管等级:
别名:
Polo-like kinase 1, PLK-1
检测方法:
ELISA
特异性:
Natural and recombinant bovine Serine/threonine-protein kinase PLK1
样品类型:
Serum, plasma, tissue homogenates, cell culture supernates and other biological fluids
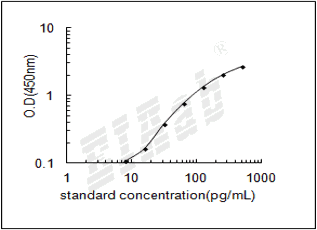
样品数据:
登录.
研究领域:
Cancer


通用注释
亚单元:
Interacts with CEP170 and EVI5. Interacts and phosphorylates ERCC6L. Interacts with FAM29A. Interacts with SLX4/BTBD12 and TTDN1. Interacts with BUB1B. Interacts (via POLO-box domain) with the phosphorylated form of BUB1, CENPU and CDC25C. Interacts with isoform 3 of SGO1. Interacts with BORA, KIF2A and AURKA. Interacts with TOPORS and CYLD. Interacts with ECT2; the interaction is stimulated upon phosphorylation of ECT2 on 'Thr-444'. Interacts with PRC1. Interacts with KIF20A/MKLP2 (when phosphorylated), leading to the recruitment at the central spindle. Interacts (via POLO box domains) with PPP1R12A/MYPT1 (when previously phosphorylated by CDK1). Part of an astrin (SPAG5)-kinastrin (SKAP) complex containing KNSTRN, SPAG5, PLK1, DYNLL1 and SGO2. Interacts with BIRC6/bruce. Interacts with CDK1-phosphorylated FRY; this interaction occurs in mitotic cells, but not in interphase cells. FRY interaction facilitates AURKA-mediated PLK1 phosphorylation. Interacts with CDK1-phosphorylated DCTN6 during mitotic prometaphase; the interaction facilitates recruitment to kinetochores. Interacts with CEP68; the interaction phosphorylates CEP68. Interacts (via POLO-box domain) with DCTN1. Interacts with FOPNL in later G1, S, G2 and M phases of the cell cycle; this interaction recruits PLK1 to centrosomes, a step required for S phase progression. Interacts with HSF1; this interaction increases upon heat shock but does not modulate neither HSF1 homotrimerization nor DNA-binding activities.
功能:
Serine/threonine-protein kinase that performs several important functions throughout M phase of the cell cycle, including the regulation of centrosome maturation and spindle assembly, the removal of cohesins from chromosome arms, the inactivation of anaphase-promoting complex/cyclosome (APC/C) inhibitors, and the regulation of mitotic exit and cytokinesis. Polo-like kinase proteins acts by binding and phosphorylating proteins are that already phosphorylated on a specific motif recognized by the POLO box domains. Phosphorylates BORA, BUB1B/BUBR1, CCNB1, CDC25C, CEP55, ECT2, ERCC6L, FBXO5/EMI1, FOXM1, KIF20A/MKLP2, CENPU, NEDD1, NINL, NPM1, NUDC, PKMYT1/MYT1, KIZ, PPP1R12A/MYPT1, PRC1, RACGAP1/CYK4, SGO1, STAG2/SA2, TEX14, TOPORS, p73/TP73, TPT1 and WEE1. Plays a key role in centrosome functions and the assembly of bipolar spindles by phosphorylating KIZ, NEDD1 and NINL. NEDD1 phosphorylation promotes subsequent targeting of the gamma-tubulin ring complex (gTuRC) to the centrosome, an important step for spindle formation. Phosphorylation of NINL component of the centrosome leads to NINL dissociation from other centrosomal proteins. Involved in mitosis exit and cytokinesis by phosphorylating CEP55, ECT2, KIF20A/MKLP2, CENPU, PRC1 and RACGAP1. Recruited at the central spindle by phosphorylating and docking PRC1 and KIF20A/MKLP2; creates its own docking sites on PRC1 and KIF20A/MKLP2 by mediating phosphorylation of sites subsequently recognized by the POLO box domains. Phosphorylates RACGAP1, thereby creating a docking site for the Rho GTP exchange factor ECT2 that is essential for the cleavage furrow formation. Promotes the central spindle recruitment of ECT2. Plays a central role in G2/M transition of mitotic cell cycle by phosphorylating CCNB1, CDC25C, FOXM1, CENPU, PKMYT1/MYT1, PPP1R12A/MYPT1 and WEE1. Part of a regulatory circuit that promotes the activation of CDK1 by phosphorylating the positive regulator CDC25C and inhibiting the negative regulators WEE1 and PKMYT1/MYT1. Also acts by mediating phosphorylation of cyclin-B1 (CCNB1) on centrosomes in prophase. Phosphorylates FOXM1, a key mitotic transcription regulator, leading to enhance FOXM1 transcriptional activity. Involved in kinetochore functions and sister chromatid cohesion by phosphorylating BUB1B/BUBR1, FBXO5/EMI1 and STAG2/SA2. PLK1 is high on non-attached kinetochores suggesting a role of PLK1 in kinetochore attachment or in spindle assembly checkpoint (SAC) regulation. Required for kinetochore localization of BUB1B. Regulates the dissociation of cohesin from chromosomes by phosphorylating cohesin subunits such as STAG2/SA2. Phosphorylates SGO1: required for spindle pole localization of isoform 3 of SGO1 and plays a role in regulating its centriole cohesion function. Mediates phosphorylation of FBXO5/EMI1, a negative regulator of the APC/C complex during prophase, leading to FBXO5/EMI1 ubiquitination and degradation by the proteasome. Acts as a negative regulator of p53 family members: phosphorylates TOPORS, leading to inhibit the sumoylation of p53/TP53 and simultaneously enhance the ubiquitination and subsequent degradation of p53/TP53. Phosphorylates the transactivation domain of the transcription factor p73/TP73, leading to inhibit p73/TP73-mediated transcriptional activation and pro-apoptotic functions. Phosphorylates BORA, and thereby promotes the degradation of BORA. Contributes to the regulation of AURKA function. Also required for recovery after DNA damage checkpoint and entry into mitosis.Phosphorylates MISP, leading to stabilization of cortical and astral microtubule attachments required for proper spindle positioning. Together with MEIKIN, acts as a regulator of kinetochore function during meiosis I: required both for mono-orientation of kinetochores on sister chromosomes and protection of centromeric cohesin from separase-mediated cleavage. Phosphorylates CEP68 and is required for its degradation. Regulates nuclear envelope breakdown during prophase by phosphorylating DCTN1 resulting in its localization in the nuclear envelope. Phosphorylates the heat shock transcription factor HSF1, promoting HSF1 nuclear translocation upon heat shock. Phosphorylates HSF1 also in the early mitotic period; this phosphorylation regulates HSF1 localization to the spindle pole, the recruitment of the SCF(BTRC) ubiquitin ligase complex induicing HSF1 degradation, and hence mitotic progression.
亚细胞位置:
Nucleus
Chromosome
Centromere
Kinetochore
Cytoplasm
Cytoskeleton
Microtubule organizing center
Centrosome
Cytoplasm
Cytoskeleton
Spindle
Midbody
localization at the centrosome starts at the G1/S transition (By similarity). During early stages of mitosis, the phosphorylated form is detected on centrosomes and kinetochores. Localizes to the outer kinetochore. Presence of SGO1 and interaction with the phosphorylated form of BUB1 is required for the kinetochore localization. Localizes onto the central spindle by phosphorylating and docking at midzone proteins KIF20A/MKLP2 and PRC1 (By similarity). Colocalizes with FRY to separating centrosomes and spindle poles from prophase to metaphase in mitosis, but not in other stages of the cell cycle (By similarity). Localization to the centrosome is required for S phase progression (By similarity). Colocalizes with HSF1 at the spindle poles during prometaphase (By similarity).
数据库链接
UniGene:
SMR:
STRING:
KEGG:
Pfam:
Uniprot:



 验证序列:
验证序列:
该产品尚未在任何出版物中被引用。
[1].
牛丝氨酸/苏氨酸蛋白激酶PLK1(PLK1)ELISA试剂盒可以做多少个样本?
牛丝氨酸/苏氨酸蛋白激酶PLK1(PLK1)ELISA试剂盒分为2种规格,96孔和48孔。96孔的试剂盒,标曲和样本都做复孔的话,可以检测40个样本。96孔的试剂盒,标曲和样本都不做复孔的话,可以检测88个样本。
[2].
牛丝氨酸/苏氨酸蛋白激酶PLK1(PLK1)ELISA试剂盒使用视频?
牛丝氨酸/苏氨酸蛋白激酶PLK1(PLK1)ELISA试剂盒实验操作视频在以下网址中,对每一步的实验步骤都做了演示,方便实验员能更好地理解ELISA实验的过程。
https://www.eiaab.com.cn/lesson-tech/805.html
https://www.eiaab.com.cn/lesson-tech/805.html
[3].
牛丝氨酸/苏氨酸蛋白激酶PLK1(PLK1)ELISA试剂盒是放在-20℃冰箱保存吗?
EIAab的牛丝氨酸/苏氨酸蛋白激酶PLK1(PLK1)ELISA试剂盒,洗涤液、底物、终止液保存于4℃,其余试剂-20℃冰箱保存。
[4].
牛丝氨酸/苏氨酸蛋白激酶PLK1(PLK1)ELISA试剂盒原理?
双抗体夹心法:用纯化的抗体包被微孔板,制成固相抗体,往包被有固相抗体的微孔中依次加入标准品或受检样本、生物素化抗体、HRP标记的亲和素,经过彻底洗涤后用底物TMB显色。用酶标仪在450nm波长下测定吸光度(OD值),计算样本浓度。
竞争法:用纯化的抗体包被微孔板,制成固相抗体,往包被有固相抗体的微孔中依次加入标准品或受检样本和生物素标记的目标分析物,受检标本中抗原与生物素标记抗原竞争结合有限的抗体。再加入HRP标记的亲和素,经过彻底洗涤后用底物TMB显色。用酶标仪在450nm波长下测定吸光度(OD值),计算样本浓度。
竞争法:用纯化的抗体包被微孔板,制成固相抗体,往包被有固相抗体的微孔中依次加入标准品或受检样本和生物素标记的目标分析物,受检标本中抗原与生物素标记抗原竞争结合有限的抗体。再加入HRP标记的亲和素,经过彻底洗涤后用底物TMB显色。用酶标仪在450nm波长下测定吸光度(OD值),计算样本浓度。
[5].
牛丝氨酸/苏氨酸蛋白激酶PLK1(PLK1)ELISA试剂盒中需要使用的样品量是多少?
夹心法100μL/孔,竞争法50μL/孔。如样本浓度过高时,应对样本进行稀释,以使稀释后的样本符合试剂盒的检测范围,计算时再乘以相应的稀释倍数。
[6].
如何分析牛丝氨酸/苏氨酸蛋白激酶PLK1(PLK1)ELISA试剂盒数据?
建议标准曲线,并计算样本浓度。对于elisa的曲线拟合,一般建议采用4参数曲线拟合,4参数曲线拟合通常更适合免疫分析。推荐使用专业软件进行曲线拟合,例如curve expert 1.3。根据样本的OD值由标曲查出相应的浓度,再乘以稀释倍数;或用标准物的浓度与OD值计算出标曲的回归方程式,将样本的OD值代入方程式,计算出样本浓度,再乘以稀释倍数,即为样本的实际浓度。以下链接是curve expert 1.3软件拟合曲线的方法。
https://www.eiaab.com.cn/news/502/
https://www.eiaab.com.cn/news/502/
[7].
牛丝氨酸/苏氨酸蛋白激酶PLK1(PLK1)ELISA试剂盒中是否包含人和动物的副产物,是否包含感染的或者传染性原料如HIV等?
除了抗体和稀释液中的BSA,不含其它人和动物的副产物,也不含感染材料。
[8].
收集牛丝氨酸/苏氨酸蛋白激酶PLK1(PLK1)ELISA试剂盒血浆样本,用什么作为抗凝剂?
一般建议用EDTA和肝素作为抗凝剂。
[9].
牛丝氨酸/苏氨酸蛋白激酶PLK1(PLK1)ELISA试剂盒酶标板可以拆成几部分?拆的时候是否需要避光,无菌?
牛丝氨酸/苏氨酸蛋白激酶PLK1(PLK1)ELISA试剂盒酶标板是8×12孔条,可拆卸,板子可以拆成12条,注意避免孔污染,不需要避光和无菌。暂时不用的板子,放回原来装的袋子里,密封保存。
[10].
牛丝氨酸/苏氨酸蛋白激酶PLK1(PLK1)ELISA试剂盒样本如何保存?
尽量检测新鲜样本。若无新鲜样本,则4℃保存1周,-20℃保存1个月,-80℃保存2个月。
反馈墙
评论数 : 0
所有用户
所有用户
默认排序
默认排序
最近
早期
目前还没有评论。




通知
规格
数量
单价 (¥)
小计 1 (¥)
小计 2:
¥

规格
数量
单价 (¥)
 折扣:
折扣:
请选择至少一个规格!